ОФИЦИАЛЬНЫЙ САЙТ ПРЕПАРАТА ПИНЕАМИН
Подтвердите, пожалуйста, являетесь ли Вы дипломированным специалистом в сфере здравоохранения?



Касян В.Н.
ГБОУ ВПО Московский государственный медико-стоматологический университет им. А.И. Евдокимова Минздрава России, Москва
Климактерический период (климакс, климактерий) — физиологический период жизни женщины, в течение которого на фоне возрастных изменений организма доминируют инволюционные процессы в репродуктивной системе. Он характеризуется постепенным снижением и «выключением» функции яичников в связи с истощением запаса яйцеклеток, и развитием симптомов эстрогенодефицита. До 60—80% женщин в пери- или постменопаузе беспокоят вазомоторные симптомы, возникновение которых начинается в переходном периоде и достигает своего пика примерно через год после наступления менопаузы (ACOG 2014, Kronenberg 1990).
Внезапное ощущение жара в верхней части тела, особенно на коже лица, шеи и груди, называют приливами. Эти эпизоды длятся примерно от 1 до 5 минут и характеризуются потливостью, гиперемией, ознобом, ощущением липкости, тревожностью и иногда учащенным сердцебиением. Вазомоторные симптомы также могут нарушать сон и являться причиной хронической бессонницы у некоторых женщин (Rance, Kronenberg). Приливы отличаются по частоте и продолжительности: исследования показывают, что у 87% женщин приливы случаются ежедневно, примерно 33% из них ощущают приливы 10 раз в день и более (ACOG). Эти симптомы значимо снижают качество жизни женщин и заставляют их обращаться за медицинской помощью.
Патофизиология приливов жара до настоящего времени полностью не раскрыта. Известно, что приливы есть у женщин, принимающих тамоксифен по поводу рака молочной железы, после овариэктомии, а также при гипогонадизме у мужчин, на фоне андроген-блокирующей терапии по поводу рака простаты (Stearns 2002; Santoro 2008). Понятно, что изменения уровней половых гормонов играют основную роль и являются причиной возникновения вазомоторных симптомов, что подтверждается их исчезновением при приеме эстрогенов. Однако снижение уровня эстрогенов и закономерное повышение уровня фолликулостимулирующего гормона (ФСГ) по механизмам обратной связи сами по себе не могут объяснить возникновение приливов, а также разнообразие тяжести и частоты симптомов, поскольку они не коррелируют с уровнями половых гормонов. Кроме того, несмотря на низкий уровень эстрогенов в постменопаузе, есть женщины с симптомами, а есть с их отсутствием. В основном обсуждается нарушение процессов терморегуляции в гипоталамусе в связи с дефицитом эстрогенов, что приводит к сужению терморегуляторной зоны и увеличению ее чувствительности к небольшим изменениям в температуре тела.
При этом еще в 1979 г. в экспериментах было показано, что приливы жара тесно приурочены к импульсам лютеинизирующего гормона (ЛГ) [Casper, Tataryn 1979](рисунок 1). Поэтому исследования последних десятилетий были сфокусированы именно на центральных механизмах регуляции репродуктивной функции.
В 1977 г группы Schally и Guillemin были удостоены Нобелевской премии за открытие гонадотропин-релизинг-гормона (ГнРГ). В 1980 г Knobil была продемонстрирована важность пульсаторной секреции ГнРГ для адекватной активации гипоталамо-гипофизарно-яичниковой оси (ГГЯ-оси) при половом созревании и ее циклической функции в репродуктивном возрасте. Высвобождение ГнРГ носит пульсовой характер, при этом оно четко коррелирует с импульсами ЛГ в периферической крови (Moenter 1992, Glanowska 2014).
Механизмы обратной связи продолжают действовать и в постменопаузе, поэтому закономерно на снижение синтеза эстрогенов яичниками повышается уровень гонадотропинов. Несмотря на четко определенную роль в период полового созревания и поддержания репродуктивной функции у взрослых, мало известно о том, как изменяется высвобождение ГнРГ в процессе старения и играет ли это роль в инволюции репродукции (Gore 2004).
Известно, что высвобождение ГнРГ у стареющих приматов, в том числе у женщин, не контролируется напрямую уровнями гонадотропинов (24-26 статья Gore). Так, в исследовании Woller (2002) у самок обезьян было показано увеличение пульсативной активности ЛГ по мере старения. У женщин уровни гонадотропинов были выше в ранней постменопаузе в сравнении с пременопаузой (27-29). Кроме того, гипофиз сам по себе может претерпевать процессы старения, к примеру, изменяя чувствительность к гонадолиберину, соответственно, синтез гонадотропинов. Поэтому важно было напрямую изучить процессы пульсативной активности ГнРГ по мере старения. В связи с недоступностью терминалей ГнРГ-нейронов и невозможностью обнаружения пептида ГнРГ в периферической крови провести такой исследование у женщин не представлялось возможным. В результате экспериментов на макаках было показано, что пульсаторная секреция ГнРГ сохранялась в различных возрастных группах, однако, для особей в постменопаузе характерны значимое повышение концентрации ГнРГ и наличие импульсов крайне большой амплитуды, при этом частота импульсов оставалась прежней (рисунок 2). В ходе эксперимента на одной обезьяне назначение эстрогенов приводило к снижению амплитуды пульсативного высвобождения ГнРГ (Gore).
В экспериментах на самках обезьян также было показано, что применение заместительной терапии эстрогенами снижало высвобождение ГнРГ (Chongthammakun) и экспрессию гена ГнРГ (El Majdoubi, Krajewski). Назначение эстрогенов женщинам в постменопаузе приводит к снижению уровней ЛГ и ФСГ, при этом ответ женщин в поздней постменопаузе был большим в сравнении с ранней постменопаузой. Это еще раз доказывает, что отрицательная обратная связь между эстрогенами и центральными регулирующими механизмами не только сохраняется, но даже укрепляется по мере старения (Gill 2002; 25, 62).
Однако временная связь между приливами и пиками ГнРГ/ЛГ не является причинной-следственной. Назначение эстрогенов и их последующая отмена все равно вызывало приливы жара у женщин даже после хирургического удаления гипофиза (Casper & Yen 1985) или на фоне агонистов гонадотропин-релизинг гормона, вызывающих подавление ЛГ (Casper & Yen 1981; DeFazio 1983). Сам по себе ГнРГ также не является причиной вазомоторных жалоб, поскольку приливы все равно имеют место у пациенток с синдромом Кальмана, несмотря на отсутствие ГнРГ-нейронов (Gambone 1984). Поскольку доказано, что импульсы ЛГ обусловлены импульсами ГнРГ, а приливы жара приурочены к импульсам ЛГ, стало понятным, что механизм возникновения приливов жара тесно связан с гипоталамическим контролем секреции ГнРГ.
Исследования начала 2000-х годов позволили доказать, что пульсативная секреция ГнРГ модулируется субпопуляцией нейронов аркуатного ядра, экспрессирующих альфа-эстрогеновый рецептор (ERα), рецептор к нейрокинину-3, кисспептин, нейрокинин В и динорфин – так называемые KNDy-нейроны (Mittelman-Smith). Одним из ярких открытий XXI века в области нейроэндокринологии было доказательство участия кисспептина и его рецептора в инициации пубертата и поддержании репродуктивной функции (Seminara&Kaiser 2005, Navarro 2013). Кисспептин является основным стимулятором синтеза гонадолиберина, его секреция также носит пульсативный характер и четко коррелирует с пульсами секреции ГнРГ/ЛГ: после каждого пульса секреции кисспептина происходит пульс секреции гонадолиберина (Navarro 2013, Guierriero 2012). Нейрокинин вместе с кисспептином оказывает стимулирующее действие на секрецию ГнРГ (Navarro 2013), динорфин – эндогенный опиоидный пептид, медиирующий отрицательную обратную связь между эстрогенами и ГнРГ (Goodman. Endocrinology. 2004; 145: 2959-2967).
Kiss1-нейроны аркуатного ядра инфундибулярной области (группа КNDy-нейронов) участвуют в отрицательной обратной связи половых стероидов на гонадотропины, обеспечивая тоническую секрецию ГнРГ, с участием динорфина (супрессора синтеза ГнРГ) и нейрокинина В (стимулятора синтеза ГнРГ). Они формируют аутосинаптическую петлю обратной связи и модулируют пульсативную секрецию ГнРГ и, соответственно, синтез ЛГ и ФСГ (Hu et al. Gen Comp Endocrinol 2014; 1; 208: 94-108). Kiss1-нейроны ростральной поля III желудочка играют ключевую роль в передаче положительной обратной связи эстрадиола на гонадотропины, в первую очередь, регулируя пульсативную активность гонадолиберина, в присутствии активированных рецепторов прогестерона, что инициирует преовуляторный пик ГнРГ [SMITH, Rance, Mittelman-Smith](рисунок 3).
В постменопаузе KNDy-нейроны претерпевают необычную соматическую гипертрофию и экспрессируют в повышенном количестве транкскрипты генов кисспептина и нейрокинина В (Mittelman-Smith, Rometo, Rance), повышается экспрессия гена kiss1 в инфундибулярном ядре, что, в свою очередь, повышает секрецию ГнРГ, ЛГ (Franceschini 2013, Alcin 2013; Rometo]. По данным исследований на животных изменения в KNDy-нейронах в постменопаузе являются вторичными по отношению к эстрогенодефициту, а не старению per se (Rometo, Rance).
При выполнении аблации KNDy-нейронов в эксперименте закономерно нарушалась секреция ЛГ. У крыс группы контроля овариэктомия закономерно приводила к повышению уровня ЛГ, а назначение эстрогенов нормализовало его. В основной группе, аблации KNDy-нейронов, после овариэктомии не происходило значимое повышение уровня ЛГ, при этом он был ниже независимо от назначения эстрогенов. Кроме того, при аблации KNDy-нейронов крысы лучше поддерживали внутреннюю температуру тела (Rance, Mittelman-Smith et al., 2012a).
Известно, что KNDy-нейроны соседствуют со структурами, отвечающими за регуляцию температуры тела (Yeo, Nakamura, Rance, Oakley), в том числе со срединным преоптическим ядром, важным компонентом термосенсорного пути защиты от перегрева. Таким образом, связи KNDy-нейронов как с преоптическими структурами, отвечающими на терморегуляцию, так и с ГнРГ-нейронами объясняет временную связь между приливами и пульсами ЛГ у женщин в постменопаузе. На основании приведенных экспериментов можно утверждать, что именно KNDy-нейроны играют важную роль в возникновении приливов жара (Rance, Mittelman).
Известно, что «золотой стандарт» купирования вазомоторных симптомов – менопаузальная гормонотерапия (МГТ), эффективность которой достигает 90-95% (метод реком МЗ, IMS 2016). В течение первых 2-3 недель приема приливы жара купируются практически полностью. По механизмам обратной связи повышение уровня эстрадиола в крови способствует уменьшению гиперактивации KNDy-нейронов, снижению амплитуды пульсовой секреции ГнРГ и уровней гонадотропинов. Прекращается передача гипервозбуждения соседним гипоталамическим структурам, ответственным за вазомоторные проявления.
Следует помнить, что у МГТ, как у любой медикаментозной терапии, есть свои показания и противопоказания. Так, МГТ противопоказана женщинам с перенесенным ранее инфарктом или инсультом, венозным тромбозом. Кроме того, с позиции баланса положительных эффектов и профиля риска, мировое сообщество рекомендует впервые назначать гормонотерапию женщинам не старше 60 лет или при длительности менопаузы не более 10 лет (метод реком МЗ, IMS 2016). Однако по данным мета-анализа шести исследований для оценки естественного развития вазомоторных симптомов было продемонстрировано сохранение приливов жара у 50% женщин через 4 года после менопаузы, а у 10% - даже через 10 лет [Politi]. Как же можно помочь пациенткам с вазомоторными жалобами, которым противопоказана МГТ или которым уже поздно ее начинать, с учетом того, что эффективность фитопрепаратов в среднем составляет около 30%, а ингибиторов обратного захвата серотонина – 50%? Существуют ли иные механизмы для устранения гиперактивации KNDy-нейронов?
2000 год ознаменовался знаковым событием в области нейроэндокринологии – открыт гонадотропин-ингибирующий гормон (ГнИГ)(Tsutsui 2000). У млекопитающих тела ГнИГ-синтезирующих нейронов располагаются в дорсомедиальной области гипоталамуса, а их аксоны подходят к срединному возвышению и к ГнРГ1-нейронам в преоптическом ядре. Гонадоингибин тормозит синтез и высвобождение гонадотропинов за счет прямого эффекта на гонадотрофы гипофиза и вызывает ингибирование активности ГнРГ-нейронов через свой рецептор, а также нарушает пульсовую секрецию ГнРГ в гипоталамусе (Ubuka 2008, 2009b)(рисунок 4). Кроме того, аксоны ГнИГ-нейронов походят к кисс-нейронам и снижают их активность. Таким образом, уровень и пульсовой ритм секреции гонадолиберина, ФСГ, ЛГ зависит от периодичности воздействия кисспептина и гонадоингибина [Wahab 2015].
По данным экспериментов на овариэктомированных мышах, при внутривенном введении ГнИГ происходило значительное снижение плазменных уровней ЛГ, а внутримозговое введение не влияло ни на средние уровни ЛГ, ни на частоту пульсативной секреции (Murakami 2008). В исследовании на культуре клеток применение ГнИГ не влияло на базальную секрецию, но снижало ГнРГ-стимулированную секрецию ЛГ в среднем на 25% (Rizwan 2009). В исследованиях было замечено, по данным некоторых авторов, имеет значение именно длительное применение ГнИГ, поскольку единичная инъекция не оказывала никакого эффекта.
По данным современных исследований, деятельность ГнИГ-нейронов модулируется как факторами внешней среды, так и внутренними факторами. Одним из таких факторов является эндогенный мелатонин. В эксперименте у птиц применение мелатонина повышает экспрессиию мРНК гонадоингибина и высвобождение самого ГнИГ в мозге (Ubuka 2005). На ГнИГ-нейронах перепелов есть подтип рецепторов к мелатонину Mel1c. Таким образом, мелатонин напрямую воздействует на ГнИГ-нейроны, повышая секрецию и высвобождение гонадоингибина. Высвобождение ГнИГ повышается в периоды коротких световых дней, когда возрастает ночная продукция мелатонина (Chowdhury 2010)(рисунок 4). Основной объем мелатонина образуется в секретирующих клетках эпифиза – пинеалоцитах.
В 2012 г в России зарегистрирован препарат Пинеамин® (Герофарм) – экстракт полипептидов эпифиза (PPG), восстанавливающий нормальную функциональную плотность пинеалоцитов, совокупность нормально функционирующих клеток эпифиза, и стимулирующий синтез эндогенного мелатонина. После внутримышечного введения препарата полипептиды PPG попадают в нижнюю полую вену, минуя портальную систему, попадают в правое сердце и далее в артериальный круг по ветвям средней и задней мозговых артерий, минуя гематоэнцефалический барьер сразу попадают внутрь пинеалоцитов через аксо-вазальные синапсы, где активируют синтез мелатонина и серотонина. Как обсуждалось выше, мелатонин стимулирует активность ГнИГ-нейронов и синтез ГнИГ, что, в свою очередь, способствует устранению гиперактивации KNDy-нейронов и устранению приливов жара.
Эффективность Пинеамина® изучалась в многоцентровом плацебо-контролируем исследовании под руководством Прилепской В.Н. и соавт. с участием 120 женщин: группа «Пинеамин 2 курса» 60 пациенток, группа «Пинеамин 1 курс» 30 пациенток, группа «плацебо» 30 пациенток. В результате исследования продемонстрирована статистически значимое снижение проявлений климактерических жалоб по шкале модифицированного менопаузального индекса Куппермана в сравнении с группой плацебо. При этом было показано, что инъекционная форма Пинеамина обладает высокой способностью удержания эффекта, что позволяет проводить повторные курсы не чаще чем 3 – 4 раза в год. Статистически значимой разницы в частоте развития нежелательных явлений по группам (Пинеамин®, плацебо) не выявлено: толщина эндометрия по данным ультразвукового исследования, состояние шейки матки не менялись, равно как и показатели системы гемостаза, биохимического анализа крови и липидного спектра.
Таким образом, несмотря на то, что новые звенья центральных отделов регуляции функции репродуктивной системы и основные патогенетические этапы возникновения вазомоторных жалоб были открыты в последние два десятилетия, уже сегодня на рынке есть препарат с инновационным механизмом, позволяющий купировать приливы жара при благоприятном профиле безопасности.
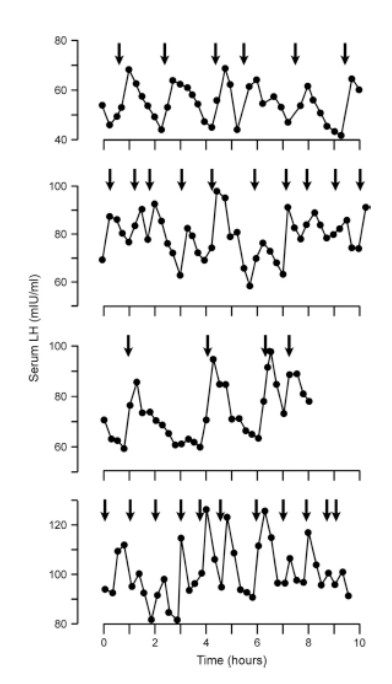
Рисунок 1. Паттерн пульсативной секреции ЛГ и эпизодов приливов жара у четырех женщин в постменопаузе. Стрелками указано начало прилива. Из статьи Casper, 1979.
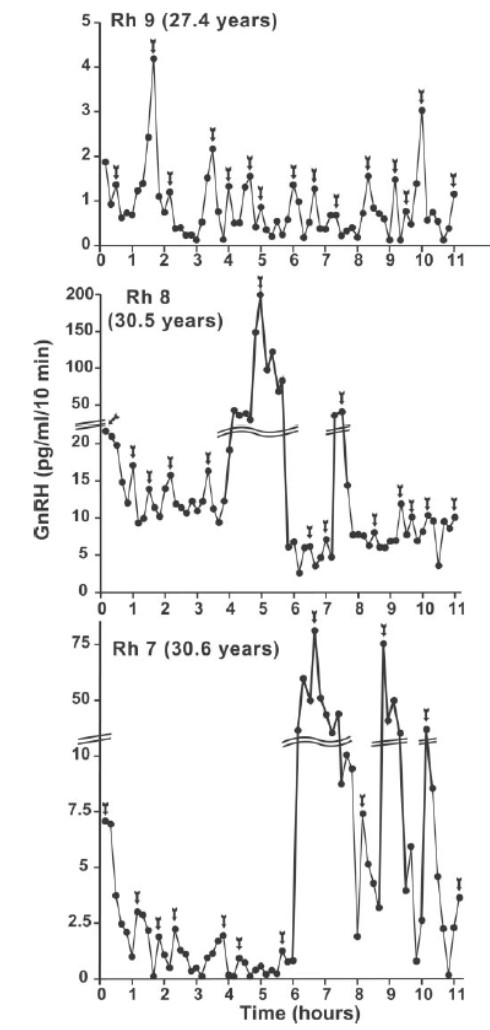
Рисунок 2. Пульсативная активность ГнРГ у самок обезьян в постменопаузе. Из статьи Gore 2004.
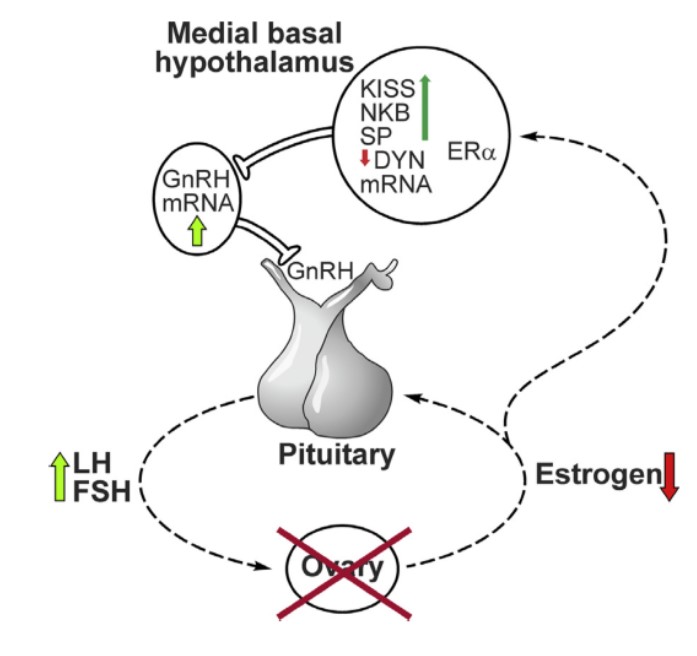
Рисунок 3. Механизм обратной связи в постменопаузе при выключении функции яичников. Из статьи Rance 2014.
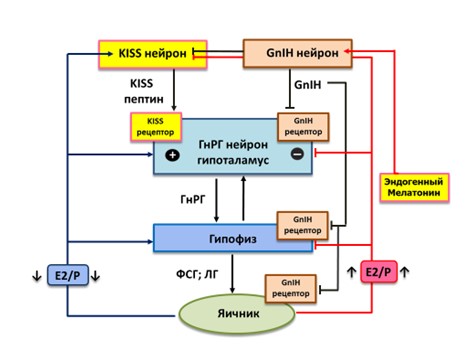
Рисунок 4. Центральные отделы регуляции репродуктивной функции. Роль мелатонина.
Подтвердите, пожалуйста, являетесь ли Вы дипломированным специалистом в сфере здравоохранения?